Z neuropsychologického hlediska můžeme lidský mozek chápat také jako stroj na kauzalitu (A » B), symetrii (oproti asymetrii), konsonanci (proti disonanci), harmonii (proti disharmonii), slast (proti ohrožení, averzi), pravidelnost (oproti nepravidelnosti) a jiné. Zdá se, že neokortex, jehož mohutným rozvojem se člověk odlišuje od většiny ostatních živočišných druhů, není ani tak výsledkem chůze po zadních a uvolnění rukou pro práci, jako spíše důsledkem narůstajícího počtu jedinců v sociálních skupinách. Z tohoto hlediska by se neokortex dal chápat jako sociální mozek, který sice souvisí s myslí a jazykem, ale prvotně zejména s rozvojem sociálních skupin.
|
|
Sociální mozek Na základě neuroanatomických a archeologických souvislostí lze předpokládat, že velikost sociálních skupin primátů a hominidů v posledních třech milionech let narůstala, přičemž tento nárůst se za posledních půl milionu let dramaticky zrychlil. Přitom se zdá, že velikost skupin je přímo úměrná velikosti „sociálního mozku“. Za sociální skupinu považujeme množinu jedinců, kteří se mezi sebou vzájemně rozpoznávají jakožto jednotlivci. U lidí lze za takovou skupinu považovat množinu jedinců, jejichž adresy máme v adresáři nebo telefonní čísla ve svém mobilním telefonu. Velikost takové skupiny je u každého člověka žijícího v současném světě zhruba 150 – 200. Sociální koheze skupin je u primátů a hominidů udržována především opečováváním, „groomingem“. Je logické, že s rostoucí velikostí skupiny také narůstá časová náročnost groomingu, který slouží jako „sociální lepidlo“. Čím větší počet jedinců je nutno opečovat v zájmu koheze skupiny, tím větší čas denní doby je třeba pro grooming alokovat. To nelze donekonečna, protože narůstající časová proporce a investice do groomingu začne vytěsňovat jiné, pro život důležité aktivity, jako je obživa, reprodukce, teritorialita, boj s přirozenými nepřáteli apod. Zdá se, že maximální únosná doba, kterou je možno beztrestně se věnovat groomingu, činí asi 30 % denní doby. Mnohé nálezy svědčí pro to, že s rostoucí velikostí skupin byla tato doba překročena zhruba před půl milionem let. To byla ve vývoji hominidů doba, kdy velikost skupiny kladla na sociální kohezi nároky, které grooming již nemohl zajistit, a vznikl tak zřejmě neobyčejný tlak na rozvoj jiného, účinnějšího „sociálního lepidla“, než je fyzické opečovávání. Máme řadu důvodů předpokládat, že to je právě doba, kdy fyzický grooming byl nahrazen groomingem „sociálním“, jenž byl zajištěn vokalizacemi a jejich modifikací (rytmus, synchronizace, melodie), které byly základem zpěvu. Od vokalizací a jejich modulací je již jen krůček k jazyku. Všimněme si, že přirozený jazyk bezděky přiznává své původní poslání jakožto sociální grooming, mluví-li o „drbech“ (drbání, klevety, rumour). Drbat ve smyslu fyzického opečovávání, jak činí primáti a jak můžeme stále ještě vidět v zoologických zahradách, a drbat ve smyslu zdánlivě neužitečné informace, jejímž hlavním cílem není řešení bezprostředního problému, nýbrž udržení sociální skupiny, jsou zřejmě dva vidy téhož. Tuto úlohu jazyka (sociální grooming) zdůrazňoval matematik a filozof Jiří Fiala, když správně poukazoval na to, že hlavním úkolem jazyka není přenos důležitých a smysluplných informací. Vyprávěl, jak byl kdysi ve Francii na návštěvě v jedné společnosti, kde prakticky nikoho neznal a přítomní si tam mezi sebou živě povídali. Když potom večer odcházel, hostitel mu třásl rukou, řka: „To jsme si krásně popovídali.“ V tomto dojmu mu nebránila skutečnost, že Jiří Fiala údajně za celý večer pronesl pouze tři slova: „Je suis d´accord“. To, čemu tedy lingvisté říkají „communion mutuel“, není vlastně nic jiného než „social grooming“.
|
|
Vnímání Naše vnímání, ať už obrazů, či zvuků, je ustrojeno tak, že zachycuje kontrasty (rozhraní) a pohyb na klidném pozadí. To je něco z vývojového hlediska nesmírně důležitého, protože zachycení pohybu protivníka, dravce, či kořisti je z hlediska přežití asi to nejdůležitější. Posadíme-li mezi ampliony nejenom malé dítě, ale třeba opici a ve stereo uspořádání přemisťujeme zdroj zvuku v prostoru, můžeme pozorovat otáčení hlavy ve směru zdroje. To je jedním z příkladů, jak vnímání pracuje na principu kontrastů, na principu odlišení signálu od šumu a popředí od pozadí. Z hlediska vývoje hudby jakožto modulace vokalizací, jež se zřejmě ve vývoji objevily jako vyšší stupeň groomingu, který může obsloužit najednou celou řadu jedinců, jsou zajímavé archeologické nálezy prvních dochovaných hudebních nástrojů. Tak kostní flétna stará zhruba 50 tisíc let z doby středního paleolitu, která byla objevena na neandrtálském tábořišti ve Slovinsku a která je zřejmě vyrobena ze stehenní kosti medvědí tlapy, má čtyři otvory. Vzdálenost mezi 2. a 3. je dvojnásobná oproti vzdálenosti mezi otvorem 3. a 4., což je v souladu s tvorbou celého tónu, resp. půltónu a naznačuje, že již tehdy měla stupnice prakticky dnešní podobu (Kunej a Turk, 2000). Základní hudební formy jsou asi předurčeny neurobiologickým substrátem, který určuje do jisté míry jejich formu. Staré čínské flétny, které jsou vlastně prvními zcela zachovanými hudebními nástroji, jež se nedávno našly v Číně a datují se mezi 7000 a 5700 let př. Kr., byly vyrobeny z kostí jeřábů a některé mají osm otvorů ve stejném uspořádání jako dnešní zobcové flétny. Na mysl, jazyk, hudbu a společnost můžeme tedy nahlížet z evolučního hlediska z jediné perspektivy. Jejich společným jmenovatelem je sociální mozek (neokortex). Shrneme-li tedy výše uvedené, pak zhruba před půl milionem let přesáhla velikost sociálních skupin primátů a hominidů zhruba stovku, což odpovídá nutnosti alokovat více než 30 % času fyzickému groomingu. V té době tedy vzniká tlak na rozvoj jiného kohezního mechanismu, kterým jsou vokalizace. Jejich modulací vzniká prototyp zpěvu resp. hudby. K tomuto rozvoji „sociálního groomingu“ dochází ruku v ruce s rozvojem sociálního mozku (neokortexu) a objevení se genu FOXP2, který je na rozvoj řeči zřejmě vázán. Evoluční stabilita pro rozvoj sociálních skupin tak důležitého sociálního groomingu je zřejmě zajištěna mechanismy libosti (prožitky krásy, slasti, radosti), kterými je opatřen.
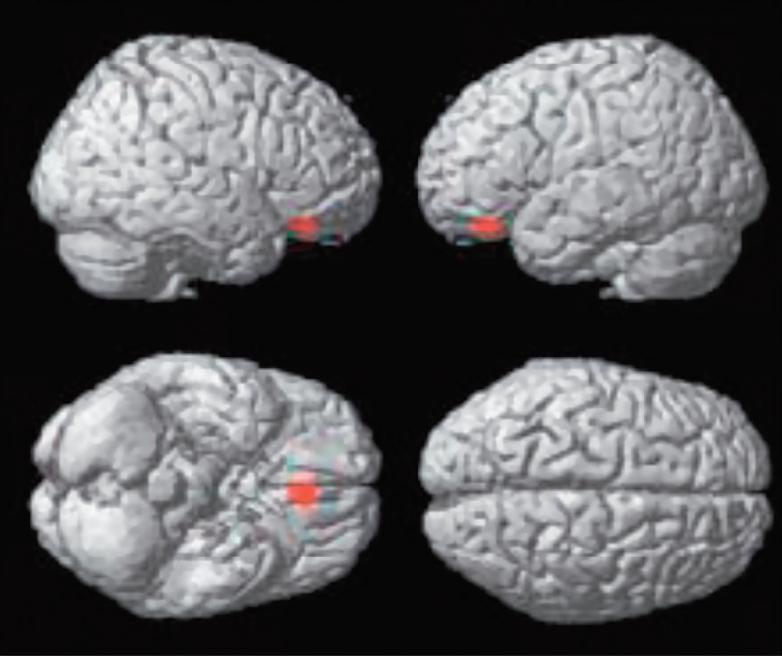
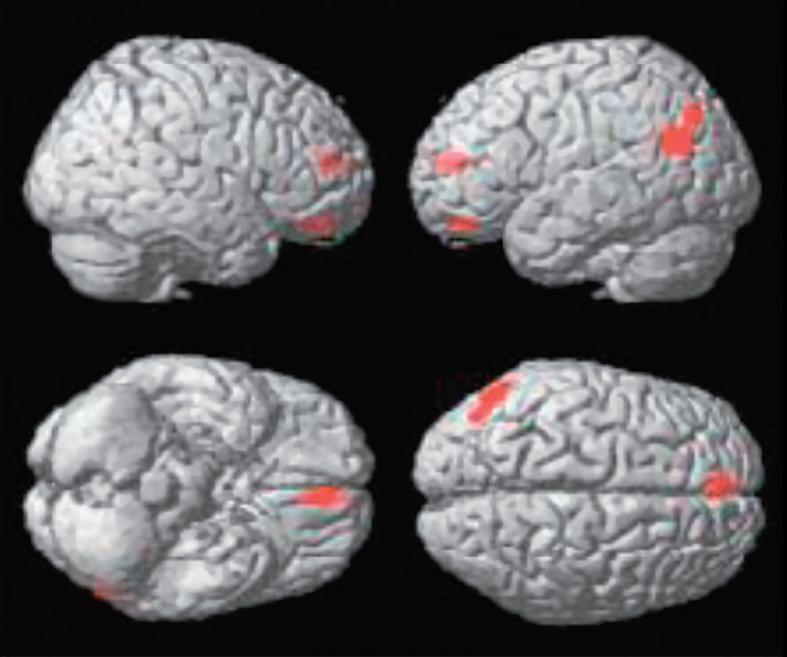
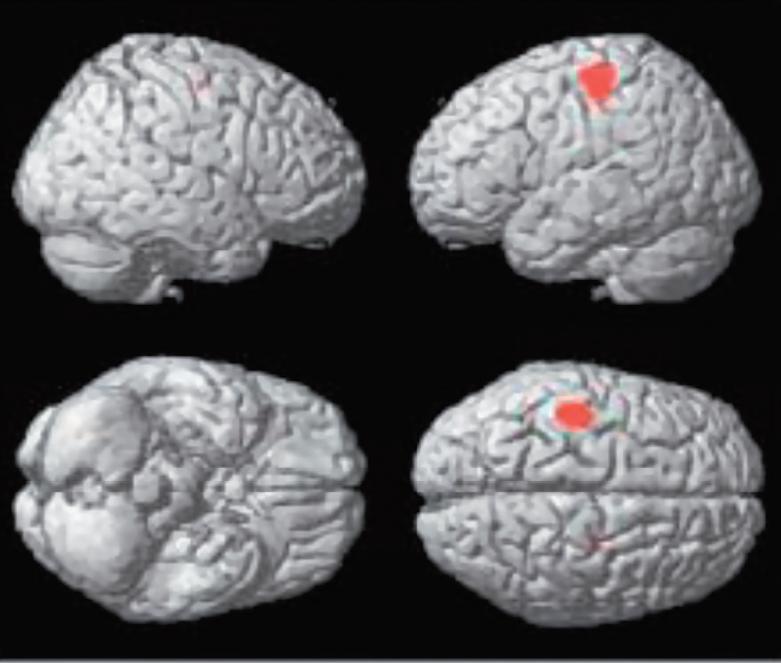
KrásaVnímání krásy je zřejmě podmíněno symetrií s přídavkem asymetrie, rytmu s trochou dysrytmie a harmonie s trochou disharmonie. Symetrie je v přírodě symbolem krásy, harmonie, a korelátem dobrého zdraví (fitness); hraje tedy v přírodním výběru důležitou roli. Absolutní symetrie má však v sobě cosi technicky sterilního. Naproti tomu výrazná asymetrie je často projevem nemoci, signalizuje hrozbu, nebezpečí, vede k úniku anebo k obraně. Přitom falešně pozitivní vyhodnocení hrozby se evolučně vyplácí daleko více než falešně negativní podhodnocení rizika. Zdá se, že libostní vyhodnocování drobných odchylek od symetrie, harmonie a rytmicity má své neuronální koreláty, které dnes lze objektivně studovat. Na obrázku, který je převzat z práce Kawabata a Zekiho (2003), je na statistických mapách, renderovaných na standardní mozek s využitím pozitronové emisní tomografie, vidět aktivita specifická pro rozhodování mezi krásou a ošklivostí, krásou a neutrálním pocitem, ošklivostí a krásou a ošklivostí a neutrálním pocitem. Pokusné osoby nejprve vyhodnocovaly na subjektivní stupnici pocity, které v nich vyvolávají obrazy, jež jim byly předloženy (ošklivý – neutrální – krásný) a posléze jim byla snímána aktivita mozku s pomocí PET za současné expozice týmiž obrazy. Přitom se nerozlišovalo, jde-li o krajinu, zátiší, portrét apod. Podobně lze dnes již objektivně detekovat neuronální základy konsonance a jejího odlišení od disonance. Je to možné při záznamu vln akustického evokovaného potenciálu snímaného z Heschlova gyru, při jehož vyhodnocení se proti sobě vynáší frekvence (Hz) a amplituda (μV). Je-li pokusná osoba nebo primát exponována konsonantním akordům (zvýšená kvarta, velká kvinta, oktáva), obě křivky běží paralelně, zatímco při expozici disonantním intervalům (malá sekunda, velká sekunda, velká septima) dochází „resonanci“, která se v elektrofyziologii označuje jako „locked in frequency“.
|
|
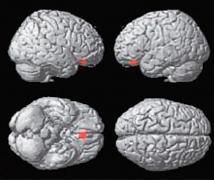 | | (A) Krásný versus ošklivý |
|
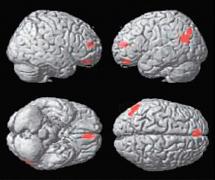 | | (B) Ošklivý vs krásný |
|
|
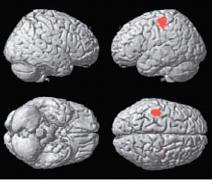 | | (Ošklivý vs neutrální) |
|
 | | (D) Krásný vs neutrální |
|
|
Statistické parametrické mapy renderované na standardní mozek, které ukazují aktivitu specifickou pro rozhodování mezi krásou a ošklivostí (A), krásou a neutrálním pocitem (B), ošklivostí a krásou (C) a ošklivostí a neutrálním pocitem (D) napříč výtvarnými tématy (korigováno, p<0.05).
(A) Pouze mediální orbito-frontální kůra, (B) mediální orbitofrontální kůra, přední gyrus cinguli a levá parietální kůra.
(C) somato-motorická kůra oboustranně, (D) žádná aktivita na korigované signifikantní úrovni.
Kawabata and Zeki, 2003
|
|
Dekonstrukce výtvarného umění a hudby ve 20. stoletíZdá se, že za posledních čtvrt milionu let vokální a grafické projevy činnosti lidského sociálního mozku nabývaly postupně na složitosti, a to až do konce 19. století. Hudba dosáhla, mj. pod vlivem církevního dogmatu, v Evropě mohutných rozměrů od Bachovského baroka přes Mozartovský klasicismus a Beethovena až po Dvořákovsko-Brahmsovský romantismus. Výtvarné umění se dostalo od gotických madon a religiózní ikonografie přes lovecká zátiší až k realistickým zobrazením bitevních výjevů. Na konci 19. století se však cosi zásadního stalo, co předznamenalo prudkou otočku ve vývoji jak hudby, tak výtvarného umění. Tu třeskutou změnu vyvolaly dva vynálezy: daguerotyp a fotografie, která vzala výtvarnému umění jeho prominentní úkol zobrazovat skutečnost, a voskový váleček a gramofon, který umožnil kopírovat hudební zážitek bez nutnosti aktivního přispění zúčastěných. Zbavena formálních pout, dekonstruovala se hudba 20. století směrem od Wagnera, přes Janáčka, Schönberga a Bouleze až k jakési atomální, „nukleární“ hudbě a podobně výtvarné umění od romantických pláten Delacroixe, přes impresionisty (Maneta), Cézanna, Braquea a Mondriana až po pop-art a Malewicze, které lze chápat jako jakési „nukleární“ výtvarné umění (viz obr. níže).
Hudba a jazyk Mnohé neurofyziologické nálezy naznačují, že Brocova oblast souvisí nejenom s řečí, ale také s hudební syntaxí. Koelsch (2000) například ukázal, které části mozku se aktivují při tzv. falešném závěru v hudbě (falešný akord na závěr skladby či hudebního motivu). Navíc i další autoři ukázali, které části mozku zpracovávají jak hudební, tak slovní syntaxi. Brown a spoluautoři (2006) pozorovali regionální aktivity různých mozkových oblastí při doplňování melodie a doplňování věty („nejlepší měsíc pro kurz španělštiny v Peru je srpen, protože v Peru je touto dobou skvělé počasí“). Při generování melodie se specificky aktivovaly oblasti BA44 a BA22, zatímco pro generování věty se specificky aktivovaly BA38, BA39/40 aj. Ostatní aktivované oblasti byly oběma úlohám společné. Brown a spoluautoři uzavírají, že hudba a jazyk jsou na úrovni senzoricko- motorických kinestetických oblastí (primární sluchová kůra a motorická kůra) zpracovávány shodně, na úrovni kombinatorního zpracování komplexních sluchových struktur (senzorická BA22 a motorická BA44 a 45) jsou zpracovány paralelně a teprve na úrovni sémantického zpracování jsou zpracovávány zvlášť. Jinými slovy „Bach mluví“. Hudební sémantika tedy vykazuje s jazykem daleko víc paralel, než kdokoli očekával. Hudba, podobně jako jazyk, je hierarchickou strukturou: elementy » slova » věty » fráze » příběhy » kompozice. Podobně hierarchicky je vystavěno i řešení problémů a používání nástrojů.
Zrcadlové buňky a hra Strukturní analýza hudebního nebo i jazykového signálu zřejmě probíhá za účasti systému tzv. zrcadlových buněk. To je ko-reprezentace intencionálních, hierarchicky uspořádaných sekvenčních motorických elementů se sluchovou informací. Zrcadlové buňky jsou buňky, které se aktivují, pozorujeme-li nějakou činnost. Je dost možné, že v zrcadlovém systému hraje důležitou úlohu Brodmannova oblast 44. Základní systém zrcadlových neuronů je fronto-parietální (MNS = Mirror Neuronal System). Fronto-parietální systém zrcadlových neuronů reprezentuje akce a úmysly druhých, a to napříč modalitami, a zapřahuje vlastní motorický systém (Iacoboni a spol. 2005). Iacoboni a spol. (1999) ukázali, jaký je rozdíl v aktivaci rostrálního parietálního kortexu a Brodmannovy oblasti BA44, pozorujeme-li pohyb prstů druhé osoby oproti kontrole. Podobně je nápadný rozdíl, pozoruje-li hru na piano ten, kdo sám na klavír umí hrát, anebo ten, kdo na klavír nikdy nehrál. Tíž autoři také ukazují pozoruhodný rozdíl v aktivaci frontálního MNS, pozoruje-li pokusná osoba uchopení hrnku v různém kontextu. Jeden naznačuje pokračující snídani (konvička s mlékem, cukřenka, pečivo, med, konev s čajem), druhý kontext naznačuje dokončenou snídani (povalená konvička od mléka, dojedené pečivo, otevřená cukřenka, prázdná konvice s čajem). První kontext vyvolává předtuchu jiného úkonu než kontext druhý. V prvém případě pokusná osoba víceméně očekává přenesení šálku k ústům, kdežto ve druhém případě odnesení šálku do dřezu na nádobí nebo do myčky. Přitom se experimentátorům podařilo vlivy kontextu odfiltrovat resp. odečíst a zmapovat pouze rozdíl v aktivaci zrcadlových buněk, které reflektují uchopení hrnku jako takové a jejichž aktivace je kontextem předurčena. Systém zrcadlových neuronů je tedy jakýmsi „interface“ mezi percepcí a akcí, který znamená automatickou a nevědomou stimulaci neuronálních struktur, jež obrážejí (a „chápou“!) akce druhých. Jde tedy o mechanismus nesmírně důležitý k predikci chování druhých, což je základní zdatnost nezbytná k přežití a její rozvoj znamená nepochybnou selekční výhodu. Hra v obojím smyslu (na slepou bábu i na hudební nástroj) je patrně způsob, jakým se tento, pro život naprosto zásadní, systém vypracovává, udržuje a trénuje. Systém zrcadlových neuronů souvisí s empatií (vcítění se do druhých), s „theory of mind“ (sociální kognice), s jazykem (!) a s odlišením Já od Ne-Já. Nahlížíme-li schizofrenii jako diskonekční syndrom resp. poruchu zpracování informací, pak není bez zajímavosti poukázat právě na to, že u schizofrenních nemocných často (i premorbidně) chybí „hravost“, nedostává se jim empatie, mají narušenou sociální kognici (a sociální přizpůsobení), schizofrenní příznaky jsou z velké části poruchami řeči (jazyka) a psychotické příznaky často zahrnují ztrátu odlišení Já od Ne-Já. Připomeňme si také Crowovo upozornění, že schizofrenie je daní, kterou platíme za rozvoj jazyka. Vnímání motorické akce, jazyk i hudba využívají společné neuronální okruhy, které zřejmě u lidí souvisejí právě se systémem zrcadlových buněk. Zdá se, že lidé rozumějí komunikačním signálům (zrakovým či sluchovým, jazykovým či hudebním) na základě pochopení motorického aktu, který představují, ba na základě pochopení úmyslu (tj. možné související motorické akce), který se za tímto aktem skrývá.
Shrnutí
Neurobiologická hypotéza rozvoje umění tedy předpokládá, že to, co určovalo rozvoj čelní mozkové kůry, byla velikost sociálních skupin. Sociální koheze je zajišťována groomingem. Soudržnost skupin hominidů větších než sto jedinců už nemohla být zajištěna groomingem „face to face“ do 20–30 % času. Přitom víc času nebylo možno dost dobře obětovat, protože by tím byly ohroženy jiné, pro život důležité činnosti. Proto se objevil efektivní způsob udržování sociální koheze v podobě vokalizací a bubnování, které lze označit jako „sociální grooming“. Od něj lze sledovat přímou linii ke zpěvu a jazyku (drby). Zvuková rytmizace (bubnování do prsou, do bubnů) funguje v sociálních skupinách hominidů též jako synchronizátor (Zeitgeber), který je důležitým nástrojem při mobilizaci proti vnějšímu nepříteli. Zhruba mezi 500–200 tisíci let př. n. l. se objevuje gen FOXP2, který je spolu s neokortexem (social brain) a sociálním groomingem předpokladem rozvoje jazyka. Hudba i jazyk zaměstnávají do značné míry shodné oblasti mozku. Hra (doslova i v přeneseném slova smyslu) aktivuje zrcadlové buňky. Zrcadlové buňky (fronto-parietální systém) umožňují predikci (co bude následovat), vcítění do druhého a předpověď chování druhého (co udělá). Predikce chování druhých je základním předpokladem přežití a znamená značnou selekční výhodu. Společným jmenovatelem systému zrcadlových buněk je Brodmanova area 44. „Zbytečné“ dětské hry, honičky, říkanky, žvatlání, drby a klevety dospělých a „jalové“ činnosti jako muzicírování, zpívání, bubnování a tancování jsou všechno činnosti, které tento pro život zcela zásadní systém zrcadlových buněk trénují a naši schopnost vcítění se do druhých a predikce jejich chování utvářejí. Umění a hra tak mají zásadní evoluční význam pro rozvoj schopností vcítit se do druhého a předpovědět jeho chování, rozpoznávat emoce druhých, udržovat sociální soudržnost, a tudíž aliance, a v neposlední řadě nahlédnout sám sebe (sebereflexe). Umění a hra tedy představují společného jmenovatele rozvoje sociální dimenze jazyka, motoriky a emotivity a z vývojového hlediska jsou vzhledem k trénování schopnosti predikce nositeli značné selekční výhody.
Literatura:Brown S, Martinez MJ, Parsons LM. Music and language side by side in the brain: a PET study of the generation of melodies and sentences. European J Neurosci, 23, 2006: 2791–2803. Dunbar RIM. The social brain: mind, language, and society in evolutionary perspective. Annu Rev Antropol 2003; 32:163-181. Iacoboni M, Woods RP, Brass M, Bekkering H, Mazziotta JC, Rizzolatti G. Cortical mechanisms of human imitation. Science 286, 1999: 2526–2528. Iacoboni M, Molnar-Szakacs I, Gallese V, Buccino G, Mazziotta JC, Rizzolatti G. Grasping the Intentions of Others with One’s Own Mirror Neuron System. PloS Biology 3, 2005:529–535. Kawabata H, Zeki S, Neural Correlates of Beauty. J Neurophysiol. 2004; 91:1699-1705,. Koelsch S. Significance of Broca’s area and ventral premotor cortex for music-syntax processing. Cortex 2006; 42:518-520. Kunej D, Turk I. New perspectives on the beginnings of music: Archeological and musicological analysis of a middle Paleolithic bone “flute.” In N. L. Wallin, B. Merker, S. Brown (Eds.): The origins of music (str. 235- 268). Cambridge, MA: MIT Press, 2000. Lewis P. Musical minds. TRENDS in Cognitive Sciences, 6, 2002; 9: 364- 366. Molnar-Szakacs I, Overy K. Music and mirror neurons: from motion to ’e’motion. SCAN, I, 2006: 235–241.
Celý článek ve formátu pdf naleznete zde.
|
|
|
 |
obsah čísla 56 |
 |
ročník 2008 |
|
témata |
|

| SANQUIS PLUS |

|
| GALERIE SANQUIS |

|
| PORADNA |

|
|